1.0 INTRODUCTION
[Top] 1.1 Overview [Top] 1.2 The Thousand Islands Ecosystem
[Top] The TIE falls within the national parks system's Central Great Lakes - St. Lawrence Precambrian Region (CPS, 1990), the Great Lakes - St. Lawrence Forest Region (Rowe, 1972), and the Lakes Simcoe - Rideau Site Region 6E, Site District 6-10 (Hills, 1959) on the Canadian side. On the American side, the westerly portion of the TIE is part of the Erie - Ontario Lowland and the easterly portion is part of the St. Lawrence Lowland physiographic unit (Muller, 1965). In Ontario most of the TIE is in Leeds County, with a small portion of Frontenac County to the west. It is a part of both Erie and Jefferson counties in New York State. These political boundaries notwithstanding, the natural landscapes (vegetation communities, faunal assemblages, and physiographical features) are virtually identical on both sides of the river. The climate of the TIE is generally warmer than that of other areas at the same latitude, largely because of the proximity to Lake Ontario. The role of the Great lakes in modifying the temperatures of the surrounding shores is one of their most fundamental effects on the climate of the area (Eichenlaub, 1979). This 'lake effect' has permitted many southern species, such as Pitch Pine, the Black Rat Snake, and Deerberry, to reach the northern limit of their ranges in the TIE. The geology of the TIE is equally unique. The Frontenac Axis, a thin arm of granite that links the Canadian Shield to the north with the Adirondack Dome to the south, is the TIE's most important landscape feature. The bedrock assemblages include syenitic and associated alkalic rocks with orthoquartzite lithologies in the central part of the TIE, quartzose sandstone in the eastern part of the TIE, and a melange of granitic gneiss and granitoid intrusive rocks in the western portion (Shilts and Kettles, 1989). Shallow till, dome-shaped rock ridges, igneous cliffs, escarpments, and clay plains are the predominant landforms, together comprising the 'knobs and flats' character of the area (Chapman and Putman, 1984; White, 1993). In the river itself, the 'knobs' become the Thousand Islands. This undulating terrain, with its inherent upland and lowland topography and north-/south-/east-/west-facing slopes, produces a large variety of moisture and temperature regimes (microclimates). The soils associated with this landscape fall into four subgroups including orthic melanic brunisol soils in the northwest (parent material: very calcareous till, outwash, and deltaic), gleyed humo-ferric podzol soils in the northeast (parent material: non-calcareous medium- and coarse-textured till, deltaic, and outwash deposits), orthic humic gleysol soils in the southeast and southwest (parent material: calcareous fine-, medium-, and coarse-textures deposits from limestone and dolomite), and gleyed gray luvisol soils on Wolfe Island (parent material: calcareous fine-textured till and lacustrine deposits from limestone) (Theberge, 1989). Altitudes within the TIE are 0-200m above mean sea level (Cadman et al, 1987). The vegetation of the TIE reflects the geography, as well as the geology and climate, of the area. Forests in the TIE comprise coniferous (1.67%), deciduous (5.65%), and mixed deciduous/coniferous (12.35%) trees (See FASTLINE Study Area, Appendix A). It is estimated that the TIE has about 1000 species of plants, including 41 rare species (CPS, 1994). The TIE is also home to 35 herptile species (Cook, 1984; Kamstra and Towle, 1991) and falls within the range of 54 mammal species (Forsyth, 1985). The TIE is a significant habitat for both breeding and migratory avifauna, with 237 species cited in a recently published checklist (CPS, 1992). The TIE, and the Frontenac Axis in general, is part of the Atlantic migration flyway (Baker, 1980). Breeding birds in the TIE constitute 164 species, and about 12% (34 species) of all the breeding species in Ontario (292 species) have their greatest concentration of breeding activity recorded in the TIE (Cadman et al, 1987). Appendix C contains species lists for trees (41 species, not including ornamentals), mammals, birds, and herptiles. An estimated 80 species of fish and over 10000 species of insects may be found in the TIE. This species diversity is a reflection of the habitat diversity resulting from a combination of northern geology (the Frontenac Axis), moderated climate (the lake effect), the microclimate phenomenon (rolling topography), and a large number of ecotonal areas (especially shorelines). This diversity makes it difficult to classify the area into detailed standard associations (Woods and Holroyd, 1975). The TIE plays several important roles in the greater ecology of eastern North America. The entire Frontenac Axis can be viewed as a wildlife corridor connecting two contemporary refugia, these being Algonquin Provincial Park to the north and Adirondack Park to the south. The Thousand Islands act as "stepping stones" and stopover points for migrating species. The St. Lawrence River does present a migration obstacle (where and when it is not frozen), as does the 401 Highway, but these pale in comparison to the barrier that Lake Ontario poses to non-volant fauna. The TIE is part of a logical route around the lake. There may even be a mild funneling effect where the Canadian Shield constricts to form the north end of the Axis. Historically, there are numerous examples of species arriving in Ontario or New York via the island stepping stones. Examples of species that recently moved north into Ontario include Gypsy Moth and the Dutch Elm Disease pathogen. On the American side, moose were extirpated from Adirondack park by 1861. Between 1935 and 1980, occasional moose had wandered back into the park from Canada (and New England), with a small population of 15-20 known to exist predominantly at the north end of the park by 1987 (Saunders, 1988). The rugged terrain has remained relatively unaffected by human
development, being largely unsuitable for farming. Cultivated
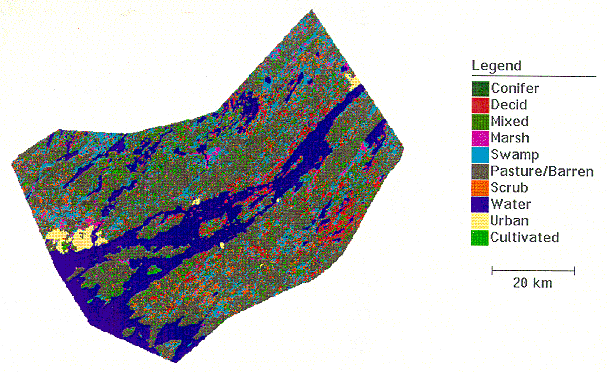
land comprises a mere 3.95% of the TIE, and urban areas make up
a meagre 1.17% of the TIE (See FASTLINE Study Area, Appendix
A). Tourism and sport-fishing are major industries in the
area. 2.0 POSTGLACIAL ECOLOGY [Top] The postglacial history of the TIE discussed in this section
spans a time period from roughly 12500 years before present (BP)
to 379 BP (1615 AD). The information comes largely from the analysis
and dating of sediment cores. These are collected from lake/river
bottoms and peat bogs. Stratigraphic analysis provides information
on floral assemblages, edaphic characteristics, fire occurrences,
and water level changes over time and can be used to infer climatic
conditions, patterns of change, and future trends. There has not
been much work of this type done in the TIE, especially on the
American side. In fact, two sediment cores (one from the St. Lawrence
River just east of Wolfe Island and one from Lambs Pond north
of Brockville) and two bog cores (one from the pitch pine ridge
on Hill Island and one from a bog near Waterton, Ontario) are
the only ones that fall within the TIE area. A core from Boyd
Pond at the north end of the Adirondacks is the only American
sample taken reasonably close to the TIE. Date estimates for events
such as warming and cooling phases and species migrations vary
considerably, but the samples do show a common general sequence
for these events. 2.1 Climate [Top] The climatic conditions and changes specific to the TIE are inferred mainly from pollen analyses of peat cores. This is based on the assumption that the presence or absence of certain species at various levels in the core sample indicates the prevailing temperature and moisture conditions at the time. Climate is one of the principal environmental controls of living organisms, with temperature being the factor of greatest biological significance (Grove, 1988). In Ontario, isotherms generally parallel latitude and vegetation zone boundaries (McAndrews, 1980), as well as the distributions of many insect species (Morgan and Morgan, 1980). The temperature changes that produce ice ages (global cooling) and interglacial periods (global warming) are thought to be the result of shifts in the Earth's orbital axis, increased/decreased sun spot activity, meteor impacts, large volcanic eruptions, the greenhouse effect, and even human activities like the burning of fossil fuels, deforestation, and chemical production (eg. CFCs causing ozone depletion). During the last Ice Age, mean global temperatures were about
5-7 degrees Celsius (ºC) cooler than today (Bradley, 1985;
GovCan, 1991; Imbrie and Imbrie, 1979). For the TIE mean January
temperatures were about -18ºC (-8ºC today) and mean
July temperatures were about +16ºC (+21ºC today) at
the last glaciation with mean annual precipitation at about 800mm
(970mm today) (McAndrews, 1980; Wright et al, 1993). The Laurentide
Ice Sheet had receded from the area by about 12500 BP with the
gradually warming temperatures (Gilbert, 1994). Since that time
there has been a period when mean global temperatures actually
increased above modern values by 1-3ºC (Anderson, 1989; McAndrews,
1980). This is referred to as the Hypsithermal Interval and occurred
between about 8000 and 4000 BP (Anderson, 1989). Significant increases
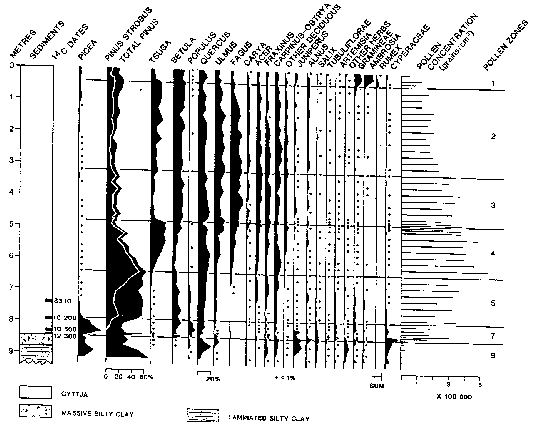
in the pollen percentages of beech, elm, and maple at about the
8000 BP mark are interpreted as the beginning of this climatic
optimum (See Appendix B). A revitalized hemlock
population around 3500 BP is indicative of the return of cooler,
moister conditions. This general decrease to normal levels prevailed
until about 750-1300 AD when another small optimum occurred (Goudie,
1983). At about 1450-1850 AD there was a marked drop in mean global
temperatures (-2ºC below modern) that is referred to as the
Little Ice Age (Bradley, 1985; Erickson, 1990; Goudie, 1983; Imbrie
and Imbrie, 1979). The Little Ice Age peaked about 1700 AD (Imbrie
and Imbrie, 1979), with a return to modern levels by about 1890
AD (Goudie, 1983). It is believed that the Little Ice Age was
caused by a 1% decrease in the sun's luminosity and the "Maunder"
Minimum of sunspot activity resulting in colder, wetter conditions
that reduced the frequency and intensity of forest fires, caused
lower snow lines and longer snow cover, slowed tree growth, forced
adjustments in the distributional ranges and migration patterns
of animals, created markedly less favourable farming conditions,
brought about a significant increase in mass movements (rockfalls,
avalanches, floods, landslides), and changed forest tree species
compositions (Erickson, 1990; Goudie, 1983; Grove, 1988). Pollen
percentages during this period show decreases in beech, elm, maple,
and especially oak (a sensitive indicator species of oncoming
ice ages), as well as a return of spruce to the TIE. It is believed
that we are presently moving towards another interglacial optimum.
2.2 Water [Top] The water history of the TIE has four distinct phases: 1) 25000-12500 BP the area was covered by the Ontario Lobe of the Laurentide Ice Sheet; 2) 12500-11400 BP the TIE was covered by glacial Lake Iroquois; 3) 11400-10500 BP the east end of the TIE was covered by the Champlain Sea, while the only water in the remainder of the TIE was a narrow estuary where early Lake Ontario drained into a southeastern arm of the Champlain Sea; and 4) 10500-present the increasing water levels of Lake Ontario were draining via the St. Lawrence River, whose levels have been controlled by the St. Lawrence Seaway lock system since 1958 AD (Anderson and Lewis, 1985; Anderson, 1989; Gilbert, 1994; Woods and Holroyd, 1975). The glacial phase resulted in a tremendous reshaping of the land. Upland areas were scoured of most or all of their surface material with the result that even today many are still essentially bare, supporting populations of mosses, grasses, and lichens or, at best, sparce forest cover (White, 1993). The weight of the ice caused isostatic depression of the Earth's crust. Maximum depression depended on the thickness and duration of the ice cover (Andrews and Peltier, 1989) and is believed to have been about 175m in the TIE area (Andrews, 1970; Fulton and Richard, 1987). Occasional deposits such as the Hill Island esker are another present-day consequence of the glacial phase. Isostatic rebound occurred most rapidly during the Lake Iroquois and Champlain Sea phases immediately following deglaciation (Gilbert, 1994). The Lake Iroquois phase produced the clay plains now found in the TIE (White, 1993). The Champlain Sea was the result of water from the east backflooding the area that was isostatically depressed. During this phase an early version of the St. Lawrence River may have been estuarine, but it is believed that the freshening influence of the glacial lakes and freshwater outflow may have been too predominant for marine flora or fauna to become established (Anderson and Lewis, 1985). At 10500 BP the river was only about 1/3-1/2 its present width (Anderson, 1989) and would not have been as significant a barrier to species migrations. The fourth phase has seen varying rates of isostatic rebound,
averaging 20 mm/a at 8000 BP, 10 mm/a at 6000 BP, and 3 mm/a at
present (Andrews, 1970). The western end of the TIE is still rising
about 1.8 mm/a with respect to the western end of Lake Ontario
(Clark and Persoage, 1970). 2.3 Soils [Top] Soil is a complex combination of minerals, air, water, and organic matter. The TIE was scoured clean by glaciation and wave-washed by Lake Iroquois, after which lake sediment accumulated in lowland areas creating clay plains. In upland areas, many of the 'knobs' described by Chapman and Putman (1984) are still barren, or at best have experienced shallow soil development, indicating that such development is indeed a very slow process. The frequency of blowdown on the islands is symptomatic of these shallow soils as trees can often only develop equally shallow root systems. Edaphic changes over time are generally described in terms
of processes such as alluviation and erosion. These processes
are directly linked to quantitative fluctuations in vegetation
cover, which in turn are often responses to climatic changes.
A postglacial sequence of these processes in North America is
described by Huggett (1991). He states that alluvial episodes
occurred roughly at 8000-6000 BP, 4500-3000 BP, and 2000-800 BP.
Before 8000 BP, vegetation change and rapid warming caused widespread
alluviation. Between 8000 BP and 7500 BP, alluviation was broken
in upon by erosion. For the next 2000 years (the Hypsithermal
Interval), warm conditions led to a retardation of alluviation.
From 6000-4500 BP, all the Holocene post-glacial valley fills
were eroded. This widespread erosive phase was the result of a
cooling climate that improved vegetation cover and reduced sediment
loads. Between about 4500 BP and 3500 BP, the rates of erosion
and deposition relaxed, only to increase again between 3000 BP
and 1800 BP. The intensity of fluvial activity then died down
again and stayed at a modest level until 1200-800 BP, when cutting
and filling, and active lateral channel migration occurred. From
800 BP to the late nineteenth century, a moderate alluviation
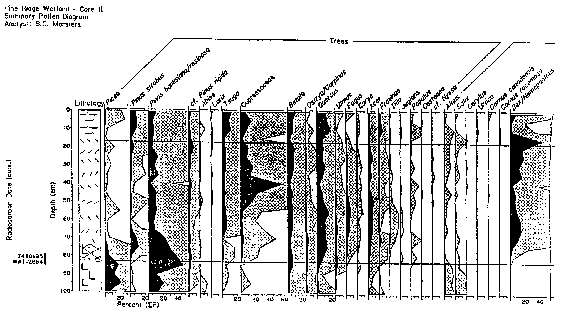
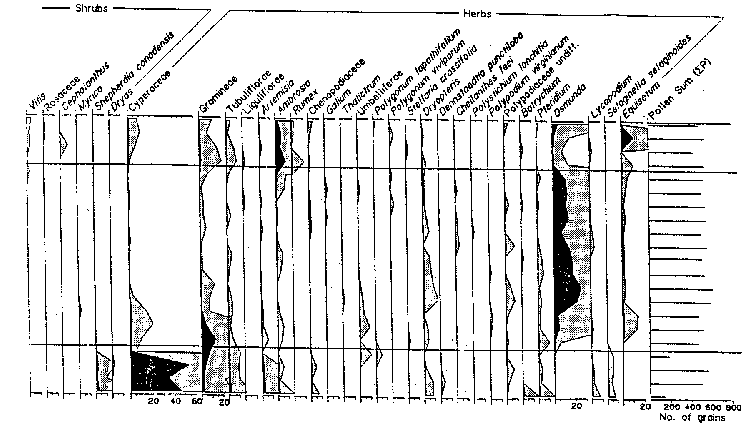
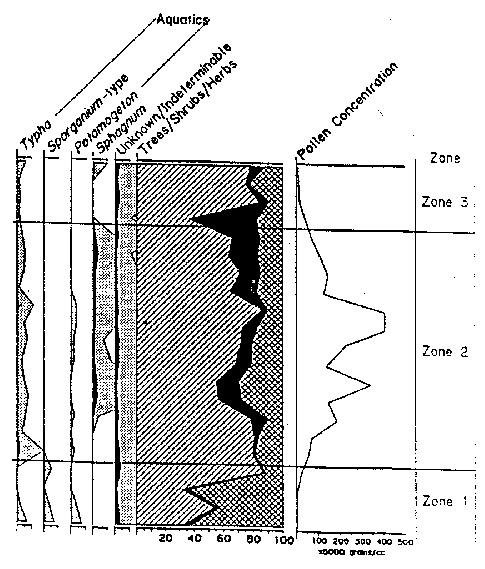
took place, after which time trenching started. 2.4 Flora [Top] Of the five core samples mentioned at the beginning of this section, only three (Lambs Pond, Boyd Pond, and Pine Ridge Bog) have had complete pollen analyses done for them (See Appendix B). The Wolfe Island core was collected and examined only for clues of past lake levels, not vegetation history. The Waterton Bog sample was unfortunately never analysed beyond the lowest levels, the information being used only to corrobate data on the Champlain Sea from the Lambs Pond sample (Anderson, 1994). It is fortuitous (with respect to this study) that Lambs Pond, Boyd Pond, and Pine Ridge Bog represent the three significant land bases of the TIE: respectively the Ontario mainland, the New York mainland, and an island. Again it should be stressed that, like climate changes, exact quantitative inferences (in this case of species compositions) are difficult to justify, but the relative sequence of events (in this case migrations) can be reasonably established. At the most general level, this sequence resembles the north-south regionality of modern vegetation zones: tundra to scrubland to boreal forest to mixedwood forest. Lake Iroquois had drained from the TIE by around 11400 BP. In the centuries preceding the complete drainage of the glacial lake, a shrub-herb tundra of grasses, willow, and wormwood, possibly intermixed with open woodlands of spruce, poplar, juniper, and alder, existed along the margins of the lake (Anderson, 1989; Gilbert, 1994). This shrub-herb assemblage was succeeded by a spruce-poplar (boreal) forest by around 10 500 BP (Anderson, 1987, 1989). The migration of spruce northward after deglaciation was faster than any other species, approximating 200km per century (Crawley, 1990). The spruce-poplar pollen zone includes boreal pines (Jack Pine, Red Pine) in the upper portion, this pine assemblage becoming the dominant forest cover by 10 200 BP. The pollen diagram for Lambs Pond shows a significant decline of both spruce and poplar pollen at this time, as well as a modest increase in oak pollen. It should be noted that all northward migrations at the end of the last ice age took place through pristine ecosystems, without the dominating influence humans that is present today (Crawley, 1990). The barrier effect of the St. Lawrence River was also not as severe as it is today in light of the lower water levels present at the time. The warming climate eventually favoured white pine growth, and a mixed pine forest with increasing amounts of white pine existed from about 8000 BP until 7000 BP. The significant increase in hemlock pollen can be attributed to hemlock growth in the shaded, cooler understory produced by the dense pine forests. The rise of hemlock coincides with a decrease in white pine, and a slower revival of birch. As well, the Hypsithermal Interval was creating warmer and moister conditions that permitted the establishment of many hardwoods, most notably elm, beech, ironwood, ash, maple, and hickory. The Hypsithermal Interval probably accelerated an inevitable migration of deciduous species leading to the mixedwood character of the TIE that still exists today. The Pine Ridge pollen diagram indicates that at 7410+/-95 BP spruce, white/red/Jack/pitch pine, hemlock, cedar, juniper, birch, ironwood, oak, elm, maple, sedges, grasses, parsley, spatterdock, wood ferns, bracken fern, cinnamon/royal fern, spike moss, cattails, and burreeds were present in the area (Warner and Marsters, 1993). The sharp decline in hemlock around 5000 BP is probably the result of a pathogenic or insectine infestation from which hemlock had recovered by about 3500-3000 BP (Anderson, 1989; Gilbert, 1994; MacDonald, 1990). No significant changes in vegetation occurred until about 500
BP. Increases in spruce, hemlock, birch, pines, and alder occur
around this time. The rise of pine, along with decreases in maple,
oak, ash, hickory, and especially beech, are often attributed
to the advent of Indian burning and land-clearing for agriculture
(Anderson, 1994; Gilbert, 1994; MacDonald, 1990; Matthews et al,
1989; Ritchie, 1987; Wright, 1994). This does not, however, explain
the increases in the other species, especially spruce. These increases
are more likely the result of the cooler climate generated by
the Little Ice Age. Burning would have promoted the growth of
pine, and led to an increase in intolerant species like birch
and poplar. Poplar does increase slightly at Lambs Pond, but more
so at Boyd Pond on the south side of the river where the Iroquois
were concentrated. However, measurable pollen counts of corn and
grasses are absent from the samples, meaning that this activity
is either over-estimated or, more likely, that agriculture was
not a very fruitful activity in the region. It is doubtful that
the natives would have had much success with their agrarian activities
in the area, given that the TIE was not an ideal environment for
farming. This aspect remains true today. The rise in ragweeds,
grasses, and dock signals the onset of intensive logging and farming
by European settlers in the 1800's (Bradley, 1985; Gilbert, 1994;
MacDonald, 1990; Matthews et al, 1989; Ritchie, 1987; Warner and
Marsters, 1993; Wright et al, 1965). This rise coincides with
reductions in most species, especially pine and hemlock, and the
arrival of many introduced plant species. On unaltered properties,
and in the region in general, the native mixedwood tree assemblages
have remained more or less the same, without extraordinary population
fluctuations. The only exceptions would be the chestnut decline
in 1880 AD and elm decline from Dutch Elm Disease since 1930 AD
(Gilbert, 1994). 2.5 Fauna [Top] It is believed that animals were crowded southward with the advancing Laurentide Ice Sheet, and returned northward from their glacial refugia in tandem with their floral associates as the ice retreated. Neither peat bog nor lake sediment cores from the TIE have yielded any significant information. There are no records of any paleofaunal finds in the TIE. This may well be because of the granitic composition of the Frontenac Axis. The limited degree of preservation of recent bone fragments found on Gordon Island is credited to the island's sandstone conglomerate (Wright, 1979) compared to the acid basic bedrock of most of the TIE. Records from across North America indicate a common pattern of major extinctions and extirpations at the start of the current interglacial interval (Huggett, 1991). In general, the large herbivores which survived were ruminants, such as moose and deer, which can live on vegetation of low diversity. They could follow their preferred food plants despite the restructuring of plant communities. Herbivores that required a broad range of food plants within their normal grazing ranges died out or were extirpated over wide areas. The demise of these large herbivores led to the extinction of the large carnivores and scavengers which preyed upon them (Huggett, 1991). Given the absence of any faunal records for the TIE in the palynology, geology, and paleoecology fields, archaeological excavation data was consulted for any clues of past faunal assemblages. The Gordon Island North Site (BbGa-2) is the only site occurring in the TIE for which faunal finds are mentioned, but the recoveries are too limited to suggest any trends through time (Wright, 1979). Species' remains identified at various stratigraphic levels included White-tailed Deer, Beaver, Black Bear, Muskrat, Raccoon, Freshwater Drum, and catfish, as well as a species of turtle and clam. A number of other sites in southern Ontario proximal to the TIE had more detailed faunal analyses done. These include sites to the west of the TIE (Auda Site, AlGo-29, near Pickering), to the north (Wyght Site, BfGa-11, Lower Rideau Lake area), and to the northeast of the TIE (Roebuck Site, near Prescott; Glenbrook Site, BgFp-5, near Cornwall; Steward Site, near Morrisburg; Beckstead Site, BfFt-1, near Prescott). Faunal finds included antlers, teeth, claws, bones, and shells. These were discussed in terms of their usage by the associated native inhabitants, and since no mention was made of possible importation of these items through trade networks it will be assumed that the identified species were locally occurring at the sites' respective time periods. Based on the latitudinal locations of the sites, and the assumption that species returned from the south as the glaciers retreated to the north, it can be inferred that the species identified at these sites were also present in the TIE at the same time or earlier. The Wyght Site, north of the TIE on the Frontenac Axis, had the earliest radiocarbon dates of all the sites. Dates ranged from 2504 BP to 874 BP (Watson, 1980). (Note: Dates in this section have been converted from "B.C." and "A.D." values to "BP" values relative to this report) Faunal remains identified to the species level included three mammals (White-tailed Deer, Beaver, and River Otter), and four fish (White Sucker, Brown Bullhead, Black Crappie, and Yellow Perch). No avifauna or herptiles were identified (Rick, 1980). The Auda Site near Pickering was dated at 1109 +/- 110 BP and revealed seven mammals (Woodchuck, Porcupine, Raccoon, Beaver, Domestic Dog, Black Bear, and a rabbit), four fish (Atlantic Salmon, Lake Trout, Whitefish, and Yellow Perch), but no identifiable birds, herptiles, or other animals (Kapches, 1987). The Roebuck Site near Prescott was dated 954 BP to 659 BP (Ross, 1994). Identified species included ten mammals (White-tailed Deer, Black Bear, Domestic Dog, Lynx, Raccoon, Beaver, Caribou, Porcupine, Woodchuck, and Moose/Wapiti), one bird (Canada Goose; a swan (probably Tundra Swan) and a loon (probably Common Loon) were also mentioned), one fish (Common Catfish), one herptile (Painted Turtle), as well as freshwater clams, a freshwater snail, and a quahog. The Steward Site near Morrisburg showed three dates that represent peak times of occupational activity: 844 BP, 609 BP, and 444 BP (Junker-Anderson, 1988). The American Eel was the only species discussed for this site. The Glenbrook Site near Cornwall was occupied around 444 BP, the faunal picture being similar to that of many sites in southern Ontario (Pendergast, 1981). Species identified include thirteen mammals (Woodchuck, Beaver, Muskrat, Porcupine, Wolf, Domestic Dog, Black Bear, Raccoon, Marten, Fisher, River Otter, White-tailed Deer, and Moose), six birds (Common Loon, Canada Goose, Bald Eagle, Sandhill Crane, Passenger Pigeon, and Common Raven), and five fish (Lake Sturgeon, Freshwater Drum, Channel Catfish, Silver Redhorse, and Shorthead Redhorse). The Beckstead Site is considered to be pre-379 BP (Ross, 1994). The large assemblage of faunal remains includes 21 mammals (Eastern Cottontail, Eastern Chipmunk, Woodchuck, Grey Squirrel, Red Squirrel, Beaver, Deer Mouse, Muskrat, Meadow Vole, Porcupine, Domestic Dog, Black Bear, Raccoon, Marten, Fisher, Mink, Striped Skunk, River Otter, White-tailed Deer, Bobcat, and Moose), five birds (Canada Goose, Ruffed Grouse, Common Tern, Passenger Pigeon, and Yellow-bellied Sapsucker), ten fish (Lake Sturgeon, Northern Pike, Longnose Sucker, Channel Catfish, Yellow Bullhead, American Eel, Largemouth Bass, Yellow Perch, Walleye, and Freshwater Drum), and one herptile (Snapping Turtle). One American report for archaeological sites in Orange County, NY, discusses early faunal finds in that area, which is in the southern part of the state. It notes radiocarbon dates of 10044+/-160 BP (a mastodon), 10994+/-150 BP (Pleistocene Moose-elk), and 12574+/-370 BP (caribou) (Funk et al, 1970). There is no evidence of the occurrence of caribou in New York during the Holocene. The above-noted species give some indication of possible fauna for the TIE at the various dates mentioned. The presence of dog remains might very well indicate an early example of a species introduction courtesy of the one mammal not discussed in section: the human being. Birds would not have been inhibited by the St. Lawrence River in their post-glacial northern migration, and would likely have returned to the TIE soon after suitable habitats had re-evolved. Mammal species from the Pickering Site may have moved around Lake Ontario via the Thousand Islands, or come from the southeastern part of the province. The diversity of habitats in the TIE, compared to the areas around the sites discussed above, may have permitted greater numbers of some species. Population fluctuations could have occurred for any number of reasons including natural disasters (eg. fire), climate changes, interspecies competition, extinctions and extirpations, diseases, or natural "boom and bust" cycles. Conclusions of any kind on the fauna of the TIE are difficult to reach, given the absence of any substantive data for the area. Some general conclusions can be drawn from all of this. Reconstructing
the changing distributions of animals and plants since the last
glaciation has brought out the transitory nature of communities.
Species abundances and distributions are constantly changing,
partly in response to climatic change. There is no "normal"
species aggregation to act as a benchmark. Huggett (1991) concluded
that biomes are temporary communities of plants and animals living
under a particular set of climatic conditions. He believed that
there is no reason to suppose that there is anything special about
the biomes we see today; that they will come and go in response
to environmental, especially climatic, changes. They are simply
the result of species adapting to the prevailing climatic regions
and gradients which have developed since deglaciation. 3.0 PRE-COLONIAL SUMMARY
[Top] The most significant ecological aspect of the pre-colonial TIE was the local ramifications of the Little Ice Age. Tree growth was slower, winters were longer and more severe, changes in faunal distribution were more substantial than normal, snow cover was prolonged, and water temperatures were lower, drastically affecting the distribution and migration of fish (Grove, 1988). The early explorers and colonists suffered the additional hardship of arriving during a period which appears to have been the coldest of the past several millenia (Eichenlaub, 1979). The natives were doubtless also affected by this climatic upheaval. Traditional hunting, gathering, and fishing patterns must have been upset by the Little Ice Age. The need to adapt to the changing faunal distributions may have resulted in the Iroquoian Wars, if one views these as an exercise in territoriality based on this need rather than bloodlust. Agricultural efforts commenced at about the same time, very likely in an effort to offset the reduced yields from the surrounding environs. If, however, farming just coincidentally started at this time, then it was an unfortunate exercise doomed to yielding poor results from the start, especially if this was being tried on the Frontenac Axis. Many animal species that are now extirpated from the TIE, or
included it in their ranges, were present in pre-colonial times.
These would include the Eastern Cougar, Elk, Moose, Grey Fox,
and Timber Wolf (Snetsinger, 1994a; Woods and Holroyd, 1975).
Water levels on the St. Lawrence River would have fluctuated more,
since there was no lock system in place. The ice would have remained
in the river longer, its availability lengthening the amount of
time that nonhibernating species had to migrate among the islands
and to the adjacent mainlands. There were also more wetland areas,
but of the same types as today. 4.0 POST-SETTLEMENT
ECOLOGY [Top] 4.1 Altered Landscapes [Top] Fires and logging destroyed much of the original forest land in the TIE (Woods and Holroyd, 1975), causing increased soil erosion and siltation that probably altered many spawning areas. These original woodlands have been mostly replaced by second- and third-growth forests of similar species composition. With regards to plants in general, introduced species now comprise over 25% of the floral assemblages on the islands alone (Woods and Holroyd, 1975). Land that was cleared for farming is still evident on larger islands, like Wolfe and Howe, as well as the clay plains of the mainland (White, 1993). Cultivated land, however, comprises only about 4% of the TIE today. Areas unsuitable for agriculture, including the many bare-rock ridges and open water, make up almost 50% of the TIE (See FASTLINE Study Area, Appendix A). Urban areas make up a mere 1.17% of the TIE, suggesting some loss of habitat. Of greater concern are the innumerable small-scale changes that have resulted from the construction of cottages, marinas, resorts, and campgrounds, as well as property modifications made by their owners (eg. replacement of natural shorelines with concrete or stone). Urbanization, tourism development, and agriculture have all contributed to habitat loss and, along with the associated infrastructures like roads and hydro corridors, have produced much fragmentation of terrestrial habitats. While this has definitely been detrimental to species requiring large areas of undisturbed wilderness, it has been a favorable development for others that prefer more open terrain (eg. coyote) or ecotonal areas. The human presence has increased the amount of pollution (sewage,
pesticides, fertilizers, landfills, litter, effluent, leachate)
in the TIE. Most of the pollution in the St. Lawrence River, however,
comes from the Great Lakes. Water levels in the river are now
artificially controlled by the lock network associated with the
St. Lawrence Seaway. 4.2 Unaltered Landscapes [Top] A preliminary investigation of possible old-growth forests in eastern Ontario was done by White in 1990. A field survey of thirteen candidate sites was conducted, including three sites within the TIE. One of these three sites, Collins Lake, situated north of Kingston, was the only site of all thirteen surveyed whose potential old-growth designation was deemed "probable". The other two sites, Weston Island and Fernbank, were allotted a "possible" designation. All three sites exemplify the deciduous/coniferous mixture characteristic of the Forest Region (See Section 1.2 and Appendix A), and have a combined area of about 40 hectares. Complete survey results for these three sites are presented in Appendix D. A number of other possible old-growth sites in the TIE that were not investigated during the study were mentioned in the report. These include Browns Bay, Camelot Island (SLINP), Charleston Lake, Gordon Island (SLINP), Hill Island (SLINP), Fitzsimmons Mountain, two sites on Grenadier Island (SLINP), Jones Creek, Leaders Creek, and South Lake. Further research may indicate similar sites on the American side. The TIE is an area with a relatively large number of wetlands
compared to the rest of southern Ontario. Approximately half of
these are marshlands, most occurring along the St. Lawrence shoreline
(Bardecki, 1984). The FASTLINE Study Area (aka TIE) shows that
marshland comprises 3.02% of the TIE (See Appendix
A). This figure does not count wetlands in the ecotonal and
shallower areas of the river, which are included in the 21.19%
of the TIE that is classed as water. Swampland in the TIE makes
up 10.88% of the total area. No percentages for bogs and fens
are given: they are included in the percentage figures for swamps
and water. Marshes and swamps are relatively short-lived wetlands,
usually created by flooding, whereas bogs and fens require more
than 170 years to develop (Snetsinger, 1994). These could be considered
old-growth. Their initial presence is attributed to the blocked
drainages resulting from various glacial deposits. The negligible
urbanization and relatively low agricultural development in the
TIE have left many of these wetlands largely undisturbed. In the
less populated northern counties of New York State many peatlands
remain as they were. Few of these are in protective ownership,
but various government agencies and private groups such as the
Nature Conservancy have established some preserves and continue
to secure other sites (Johnson, 1985). Not only are they "vaults
of history" for palynologists and other scientists, they
are vital floral and faunal habitats that represent an important
element of the TIE's biodiversity. 5.0 IN THE FUTURE
[Top] Within the TIE, a number of current trends may manifest themselves within the next 25 years. As a "playground" for people, there may be increased demand and use of the area by nearby urban populations, bringing with it added stress on the landscape. Increasing urbanization of rural areas, and pressures to cultivate marginal farmlands would further reduce available habitat (eg. forest clearing, wetland draining) and, consequently, biodiversity (Crawley, 1990). Planning reforms in Ontario are pointing towards increased development control at the municipal level, which may result in increased habitat fragmentation and less consistent environmental protection. On the agricultural front, there is a definite trend towards decreased activity within the TIE. From 1981 to 1991 Leeds County experienced a 25% drop in the number of census farms and an 18% decrease in the total area of census farms (OMAF, 1994). Old-field succession, a term for the gradual change from grasses to weeds to shrubs to forest specific to abandoned farmlands, will increase proportionately if these areas remain abandoned. Farmland that continues to show economic viability will probably not change (Snetsinger, 1994a). The TIE's second- and third-growth forests will continue to tend towards self-perpetuating climax communities with an increasing number of deciduous species as mean temperatures continue to increase. At present rates of climatic change, a 1ºC increase in mean global temperatures within the next 25-50 years will bring about significant changes in species assemblages and distributions. For the TIE, this could mean the extirpation of boreal tree species (eg. Striped Maple), and increases in Carolinian tree species (eg. hickories). There will also likely be an increase in the rate of establishment of alien flora, and an increase in the rate of spread of currently established aliens (Crawley, 1990). Surviving native species may be less vigorous following such a climatic change, increasing their susceptibility to insect and disease organisms, which will be able to adapt quicker because of faster generational turnover. Another trend has been the recognition of our remaining natural
areas as vital towards the safeguarding of biodiversity. Environmental
management is tending away from specific issues, leaning more
towards holistic ecosystem approaches. There is an increasing
network of heritage protection agencies and properties through
the formation of partnerships and conservancies, as well as continued
research into old-growth communities, Areas of Natural and Scientific
Interest (ANSI's), and many aspects of ecology, leading to a better
understanding of the natural world around us. For the TIE, and
natural areas in general, there is cause for some measure of optimism.
Anderson, T.W., and C.F.M.
Lewis. "Postglacial Water-level History
of the Lake Ontario Basin" in Quaternary Evolution of
the Great Lakes (P.F. Karrow, P.E. Calkin,Eds.). Geological
Association of Canada Special Paper 30, 1985. Wright, H.E. et al (eds.). "Vegetation, Lake Levels, and Climate
in Eastern North America for the Past 18,000 Years" and "Climatic
Changes during the Past 18,000 Years: Regional Syntheses, Mechanisms,
and Causes" in Global Climates since the Last Glacial
Maximum. Minneapolis, Minnesota: University of Minnesota Press,
1993: 415-467, 514-532. APPENDIX A [Top] FASTLINE Study Area
LAMB's POND, ONTARIO BOYD POND, NEW YORK HILL ISLAND, ONTARIO This appendix contains the common names of the native trees, herptiles, mammals, and birds whose range includes the TIE. Some are extirpated(E). An asterisk(*) denotes species that have been observed by the author since 1988. These lists can also be viewed as detailed trilingual species checklists.
PLANT LIST - SITE STATUS: C (Common) S (Sparse) R (Rare) Fraxinus americana (White Ash)............................S Fagus grandifolia (Beech)......................................C Acer saccharum (Sugar Maple)..............................C Betula papyrifera (White Birch)..............................S Tsuga canadensis (Hemlock)..................................C Prunus serotina (Black Cherry)...............................S Tilia americana (Basswood)....................................S Betula alleghaniensis (Yellow Birch)........................S Pinus strobus (White Pine).......................................R Fraxinus nigra (Black Ash)......................................R Acer rubrum (Red Maple)........................................R Cornus rugosa (Round-leaved Dogwood).................R Polystichum acrostichoides (Christmas Fern)...........C FAUNAL OBSERVATIONS: Deer, Raccoon, Black-capped Chickadee,
Porcupine,
SUMMARY 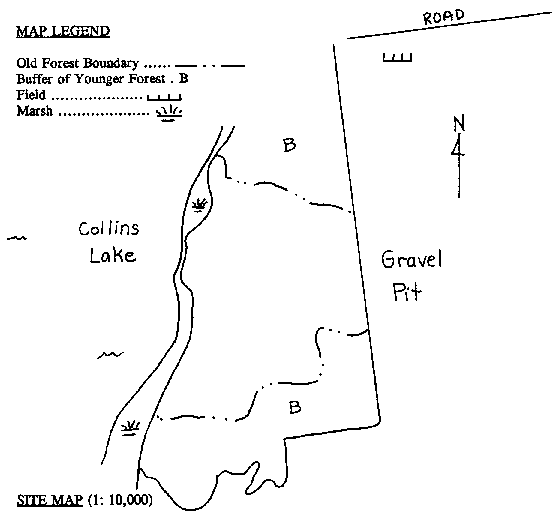
PLANT LIST - SITE STATUS: C (Common) S (Sparse) R (Rare) Pinus strobus (White Pine)........................................C Acer saccharum (Sugar Maple..................................C Fraxinus americana (White Ash)..............................C Prunus serotina (Black Cherry).................................R Quercus rubra (Red Oak)..........................................S Tilia americana (Basswood)......................................C Betula papyrifera (White Birch)..................................S Populus grandidentata (Large-leaved Aspen)............R Quercus alba (White Oak...........................................S Ostrya virginiana (Ironwood).....................................S Betula allegheniensis (Yellow Birch)...........................R Fagus grandifolia (Beech)..........................................S Acer rubrum (Red Maple)..........................................R Prunus virginiana (Choke Cherry).............................S Pteridium aquilinum (Braken Fern)............................C FAUNAL OBSERVATIONS: White-breasted Nuthatch, Brown
Creeper, Hairy
SUMMARY 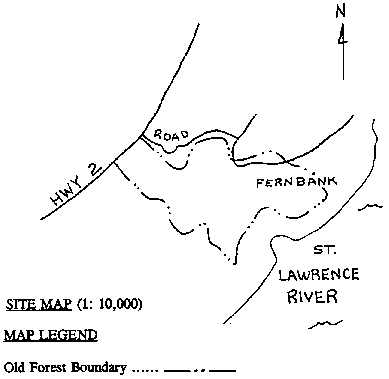
REMARKS: Species composition varies from area to area
within stand. Hamamelis virginiana (Witch Hazel).............................C Polypodium virginianum (Common Polypody)..............S FAUNAL OBSERVATIONS: Raccoon, Porcupine, Brown Creeper
SUMMARY  This appendix contains names, addresses and/or phone numbers of people and organizations that contributed information to this study. They provided published and/or unpublished documents, and where there was an absence of actual research they offered personal insights that were particularly useful. Suggestions for additional individuals or organizations to contact (*) were plentiful but the time constraints of this project, and an inability to contact some of them, left these possible sources unexplored. They are included here for anyone wishing to pursue aspects of this report further. Dr. Thane W. Anderson, Palynologist Carleton University Don Cuddy, Natural Heritage
Ecologist *Dr. William Engelbrecht, Professor
Geological Survey of Canada |